

Энергия в мышцах освобождается при. Механизм мышечных сокращений. Функции и свойства скелетных мышц. Одиночное и тетанические сокращения
Энергия в мышцах освобождается при. Механизм мышечных сокращений. Функции и свойства скелетных мышц. Одиночное и тетанические сокращения
Мышечное сокращение является жизненно важной функцией организма, связанной с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами. Все виды произвольных движений – ходьба, мимика, движения глазных яблок, глотание, дыхание и т. п. осуществляются за счет скелетных мышц. Непроизвольные движения (кроме сокращения сердца) – перистальтика желудка и кишечника, изменение тонуса кровеносных сосудов, поддержание тонуса мочевого пузыря – обусловлены сокращением гладких мышц. Работа сердца обеспечивается сокращением сердечной мускулатуры.
Структурная организация скелетной мышцы
Мышечное волокно и миофибрилла (рис. 1). Скелетная мышца состоит из множества мышечных волокон, имеющих точки прикрепления к костям и расположенных параллельно друг другу. Каждое мышечное волокно (миоцит) включает множество субъединиц – миофибрилл , которые построены из повторяющихся в продольном направлении блоков (саркомеров). Саркомер является функциональной единицей сократительного аппарата скелетной мышцы. Миофибриллы в мышечном волокне лежат таким образом, что расположение саркомеров в них совпадает. Это создает картину поперечной исчерченности.

Рис. 1. Иерархические уровни организации скелетной мышцы (по Bloom , 1968).
Саркомер и филламенты. Саркомеры в миофибрилле отделены друг от друга Z -пластинками, которые содержат белок бета-актинин. В обоих направлениях от Z -пластинки отходят тонкие актиновые филламенты. В промежутках между ними располагаются более толстые миозиновые филламенты .
Актиновый филламент внешне напоминает две нитки бус, закрученные в двойную спираль, где каждая бусина – молекула белка актина . В углублениях актиновых спиралей на равном расстоянии друг от друга лежат молекулы белка тропонина , соединенные с нитевидными молекулами белка тропомиозина.
Миозиновые филламенты образованы повторяющимися молекулами белка миозина . Каждая молекула миозина имеет головку и хвост . Головка миозина может связываться с молекулой актина, образуя так называемый поперечный мостик .
Клеточная мембрана мышечного волокна образует инвагинации (поперечные трубочки), которые выполняют функцию проведения возбуждения к мембране саркоплазматического ретикулума. Саркоплазматичекий ретикулум (продольные трубочки) представляет собой внутриклеточную сеть замкнутых трубочек и выполняет функцию депонирования ионов Са++ .
Двигательная единица. Функциональной единицей скелетной мышцы является двигательная единица (ДЕ). ДЕ – совокупность мышечных волокон, которые иннервируются отростками одного мотонейрона. Возбуждение и сокращение волокон, входящих в состав одной ДЕ, происходит одновременно (при возбуждении соответствующего мотонейрона). Отдельные ДЕ могут возбуждаться и сокращаться независимо друг от друга.
Молекулярные механизмы сокращения скелетной мышцы
Согласно теории скольжения нитей, мышечное сокращение происходит благодаря скользящему движению актиновых и миозиновых филламентов друг относительно друга. Механизм скольжения нитей включает несколько последовательных событий.
Головки миозина присоединяются к центрам связывания актинового филламента (рис. 2, А).
Взаимодействие миозина с актином приводит к конформационным перестройкам молекулы миозина. Головки приобретают АТФазную активность и поворачиваются на 120 ° . За счет поворота головок нити актина и миозина передвигаются на «один шаг» друг относительно друга (рис. 2, Б).
Рассоединение актина и миозина и восстановление конформации головки происходит в результате присоединения к головке миозина молекулы АТФ и ее гидролиза в присутствии Са++ (рис. 2, В).
Цикл «связывание – изменение конформации – рассоединение – восстановление конформации» происходит много раз, в результате чего актиновые и миозиновые филламенты смещаются друг относительно друга, Z -диски саркомеров сближаются и миофибрилла укорачивается (рис. 2, Г).

Рис. 2. Механизм мышечного сокращения.
Объяснение – в тексте.
1 – актиновый филламент,
2 – центр связывания,
3 – миозиновый филламент,
4 – головка миозина,
5 – Z -диск саркомера.
Сопряжение возбуждения и сокращения в скелетной мышце
В состоянии покоя скольжения нитей в миофибрилле не происходит, так как центры связывания на поверхности актина закрыты молекулами белка тропомиозина (рис. 3, А, Б). Возбуждение (деполяризация) миофибриллы и собственно мышечное сокращение связаны с процессом элетромеханического сопряжения, который включает ряд последовательных событий.

Рис. 3. Механизм сопряжения возбуждения и сокращения.
Объяснение – в тексте.
1 – поперечная трубочка саркоплазматичекой мембраны,
2 –саркоплазматичекий ретикулум,
3 – ион Са++ ,
4 – молекула тропонина,
5 – молекула тропомиозина.
В результате срабатывания нейромышечного синапса на постсинаптической мембране возникает ВПСП, который генерирует развитие потенциала действия в области, окружающей постсинаптическую мембрану.
Возбуждение (потенциал действия) распространяется по мембране миофибриллы и за счет системы поперечных трубочек достигает саркоплазматического ретикулума. Деполяризации мембраны саркоплазматического ретикулума приводит к открытию в ней Са++ -каналов, через которые в саркоплазму выходят ионы Са++ (рис. 3, В).
Ионы Са++ связываются с белком тропонином. Тропонин изменяет свою конформацию и смещает молекулы белка тропомиозина, которые закрывали центры связывания актина (рис. 3, Г).
К открывшимся центрам связывания присоединяются головки миозина, и начинается процесс сокращения (рис. 3, Д).
Для развития указанных процессов требуется некоторый период времени (10–20 мс). Время от момента возбуждения мышечного волокна (мышцы) до начала ее сокращения называют латентным периодом сокращения .
Расслабление скелетной мышцы
Расслабление мышцы вызывается обратным переносом ионов Са++ посредством кальциевого насоса в каналы саркоплазматического ретикулума. По мере удаления Са++ из цитоплазмы открытых центров связывания становится все меньше и в конце концов актиновые и миозиновые филламенты полностью рассоединяются; наступает расслабление мышцы.
Контрактурой называют стойкое длительное сокращение мышцы, сохраняющееся после прекращения действия раздражителя. Кратковременная контрактура может развиваться после тетанического сокращения в результате накопления в саркоплазме большого количества Са++ ; длительная (иногда необратимая) контрактура может возникать в результате отравления ядами, нарушений метаболизма.
Фазы и режимы сокращения скелетной мышцы
Фазы мышечного сокращения
При раздражении скелетной мышцы одиночным импульсом электрического тока сверхпороговой силы возникает одиночное мышечное сокращение, в котором различают 3 фазы (рис. 4, А):
латентный (скрытый) период сокращения (около 10 мс), во время которого развивается потенциал действия и протекают процессы электромеханического сопряжения; возбудимость мышцы во время одиночного сокращения изменяется в соответствии с фазами потенциала действия;
фаза укорочения (около 50 мс);
фаза расслабления (около 50 мс).

Рис. 4. Характеристика одиночного мышечного сокращения. Происхождение зубчатого и гладкого тетануса.
Б – фазы и периоды иышечного сокращения,
Б – режимы мышечного сокращения, возникающие при разной частоте стимуляции мышцы.
Изменение длины мышцы показано синим цветом, потенциал действия в мышце – красным, возбудиумость мышцы – фиолетовым.
Режимы мышечного сокращения
В естественных условиях в организме одиночного мышечного сокращения не наблюдается, так как по двигательным нервам, иннервирующим мышцу, идут серии потенциалов действия. В зависимости от частоты приходящих к мышце нервных импульсов мышца может сокращаться в одном из трех режимов (рис. 4, Б).
Одиночные мышечные сокращения возникают при низкой частоте электрических импульсов. Если очередной импульс приходит в мышцу после завершения фазы расслабления, возникает серия последовательных одиночных сокращений.
При более высокой частоте импульсов очередной импульс может совпасть с фазой расслабления предыдущего цикла сокращения. Амплитуда сокращений будет суммироваться, возникнет зубчатый тетанус – длительное сокращение, прерываемое периодами неполного расслабления мышцы.
При дальнейшем увеличении частоты импульсов каждый следующий импульс будет действовать на мышцу во время фазы укорочения, в результате чего возникнет гладкий тетанус – длительное сокращение, не прерываемое периодами расслабления.
Оптимум и пессимум частоты
Амплитуда тетанического сокращения зависит от частоты импульсов, раздражающих мышцу. Оптимумом частоты называют такую частоту раздражающих импульсов, при которой каждый последующий импульс совпадает с фазой повышенной возбудимости (рис. 4, A) и соответственно вызывает тетанус наибольшей амплитуды. Пессимумом частоты называют более высокую частоту раздражения, при которой каждый последующий импульс тока попадает в фазу рефрактерности (рис. 4, A), в результате чего амплитуда тетануса значительно уменьшается.
Работа скелетной мышцы
Сила сокращения скелетной мышцы определяется 2 факторами:
числом ДЕ, участвующих в сокращении;
частотой сокращения мышечных волокон.
Работа скелетной мышцы совершается за счет согласованного изменения тонуса (напряжения) и длины мышцы во время сокращения.
Виды работы скелетной мышцы:
динамическая преодолевающая работа совершается, когда мышца, сокращаясь, перемещает тело или его части в пространстве;
статическая (удерживающая) работа выполняется, если благодаря сокращению мышцы части тела сохраняются в определенном положении;
динамическая уступающая работа совершается, если мышца функционирует, но при этом растягивается, так как совершаемого ею усилия недостаточно, чтобы переместить или удержать части тела.
Во время выполнения работы мышца может сокращаться:
изотонически – мышца укорачивается при постоянном напряжении (внешней нагрузке); изотоническое сокращение воспроизводится только в эксперименте;
изометричеки – напряжение мышцы возрастает, а ее длина не изменяется; мышца сокращается изометрически при совершении статической работы;
ауксотонически – напряжение мышцы изменяется по мере ее укорочения; ауксотоническое сокращение выполняется при динамической преодолевающей работе.
Правило средних нагрузок – мышца может совершить максимальную работу при средних нагрузках.
Утомление – физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется снижением амплитуды сокращений, удлинением латентного периода сокращения и фазы расслабления. Причинами утомления являются: истощение запаса АТФ, накопление в мышце продуктов метаболизма. Утомляемость мышцы при ритмической работе меньше, чем утомляемость синапсов. Поэтому при совершении организмом мышечной работы утомление первоначально развивается на уровне синапсов ЦНС и нейро-мышечных синапсов.
Структурная организация и сокращение гладких мышц
Структурная организация . Гладкая мышца состоит из одиночных клеток веретенообразной формы (миоцитов), которые располагаются в мышце более или менее хаотично. Сократительные филламенты расположены нерегулярно, вследствие чего отсутствует поперечная исчерченность мышцы.
Механизм сокращения аналогичен таковому в скелетной мышце, но скорость скольжения филламентов и скорость гидролиза АТФ в 100–1000 раз ниже, чем в скелетной мускулатуре.
Механизм сопряжения возбуждения и сокращения . При возбуждении клетки Cа++ поступает в цитоплазму миоцита не только из саркоплазматичекого ретикулума, но и из межклеточного пространства. Ионы Cа++ при участии белка кальмодулина активируют фермент (киназу миозина), который переносит фосфатную группу с АТФ на миозин. Головки фосфорилированного миозина приобретают способность присоединяться к актиновым филламентам.
Сокращение и расслабление гладких мышц . Скорость удаления ионов Са++ из саркоплазмы значительно меньше, чем в скелетной мышце, вследствие чего расслабление происходит очень медленно. Гладкие мышцы совершают длительные тонические сокращения и медленные ритмические движения. Вследствие невысокой интенсивности гидролиза АТФ гладкие мышцы оптимально приспособлены для длительного сокращения, не приводящего к утомлению и большим энергозатратам.
Физиологические свойства мышц
Общими физиологическими свойствами скелетных и гладких мышц являются возбудимость и сократимость . Сравнительная характеристика скелетных и гладких мышц приведена в табл. 6.1. Физиологические свойства и особенности сердечной мускулатуры рассматриваются в разделе «Физиологические механизмы гомеостаза».
Таблица 7.1.Сравнительная характеристика скелетных и гладких мышц
Механизм мышечных сокращений кратко
Процессы мышечной работы представляют собой многоуровневый комплекс физиологических и биохимических функций, жизненно важных для полноценной работы человеческого организма. Внешне подобные процессы можно наблюдать на примерах произвольных движений при ходьбе, беге, изменении мимики и т. д. Однако они охватывают гораздо больший спектр функций, в числе которых также значится работа дыхательного аппарата, органов пищеварения и выделительной системы. В каждом случае механизм мышечных сокращений подкрепляется работой миллионов клеток, в которой задействуются химические элементы и физические волокна.
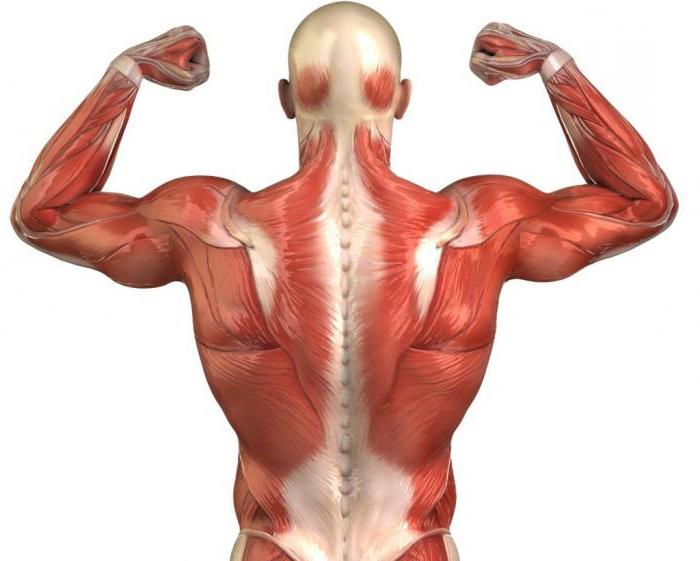
Структурная организация мышцы
Мышцы формируются множеством волокон ткани, которые имеют узлы крепления к костям скелета. Они располагаются параллельно и в процессе мышечной работы взаимодействуют между собой. Именно волокна при поступлении импульсов обеспечивают механизм мышечного сокращения. Кратко структуру мышцы можно представить как систему, состоящую из молекул саркомер и миофибрилла. Важно понимать, что каждое мышечное волокно образуется множеством субъединиц миофибрилл, располагающихся продольно по отношению друг к другу. Теперь стоит отдельно рассмотреть саркомеры и филаменты. Поскольку они играют важную роль в двигательных процессах.
Саркомеры и филаменты
Саркомеры представляют собой сегменты волокон, которые отделяются так называемыми Z-пластинами, содержащими бета-актинин. От каждой пластины отходят актиновые филаменты, а промежутки заполняются толстыми миозиновыми аналогами. Актиновые элементы, в свою очередь, похожи на ниточки бус, закрученных в двойную спираль. В этой структуре каждая бусинка является молекулой актина, а в участках с углублениями в спирали находятся молекулы тропонина. Каждая из этих структурных единиц формирует механизм сокращения и расслабления мышечного волокна, связываясь друг с другом. Ключевую роль в возбуждении волокон играет клеточная мембрана. В ней заключены поперечные трубочки-инвагинации, которые активизируют функцию саркоплазматического ретикулума – это и будет возбуждающий эффект для мышечной ткани.
Двигательная единица
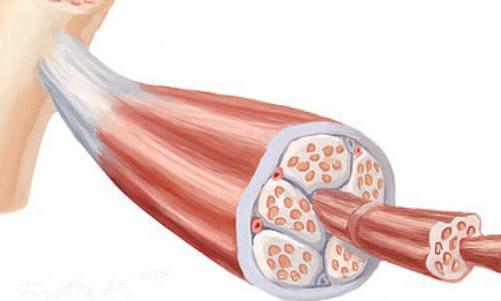
Теперь стоит отойти от углубленной структуры мышцы и рассмотреть двигательную единицу в общей конфигурации скелетной мышцы. Это будет совокупность мышечных волокон, иннервируемых отростками мотонейрона. Работа ткани мышцы независимо от характера действия будет обеспечиваться волокнами, включенными в состав одной двигательной единицы. То есть при возбуждении мотонейрона срабатывает механизм мышечных сокращений в рамках одного комплекса с иннервируемыми отростками. Такое разделение на мотонейроны позволяет целенаправленно задействовать конкретные мышцы, не возбуждая без надобности соседние двигательные единицы. По сути, вся мышечная группа одного организма делится на сегменты мотонейронов, которые могут объединяться в работе над сокращением или расслаблением, а могут действовать разнопланово или поочередно. Главное, что они независимы друг от друга и работают только с сигналами своей группы волокон.
Молекулярные механизмы мышечной работы
В соответствии с молекулярной концепцией о скольжении нитей, работа мышечной группы и, в частности, ее сокращение реализуется в ходе скользящего действия миозинов и актинов. Реализуется сложный механизм взаимодействия этих нитей, в котором можно выделить несколько процессов:
- Центральная часть миозиновой нити соединяются со связками актинов.
- Достигнутый контакт актина с миозином способствует конформационному перемещению молекул последнего. Головки вступают в фазу активности и разворачиваются. Таким образом осуществляются молекулярные механизмы мышечного сокращения на фоне перестройки нитей активных элементов по отношению друг к другу.
- Затем происходит взаимное расхождение миозинов и актинов с последующим восстановлением головной части последних.
Весь цикл выполняется несколько раз, в результате чего происходит смещение вышеупомянутых нитей, а Z-сегменты саркомеров сближаются и укорачиваются.
Физиологические свойства работы мышц
Среди основных физиологических свойств мышечной работы выделяют сократимость и возбудимость. Эти качества, в свою очередь, обуславливаются проводимостью волокон, пластичностью и свойством автоматии. Что касается проводимости, то она обеспечивает распространение процесса возбудимости между миоцитами по нексусам – это специальные электропроводящие контуры, отвечающие за проведение импульса сокращения мышцы. Однако после сокращения или расслабления тоже совершается работа волокон.
За их спокойное состояние в определенной форме отвечает пластичность, определяющая сохранение постоянного тонуса, в котором на текущий момент находится механизм мышечного сокращения. Физиология пластичности может проявляться как в виде сохранения укороченного состояния волокон, так и в их растянутом виде. Интересно и свойство автоматии. Она определяет способность мышц входить в рабочую фазу без подключения нервной системы. То есть миоциты самостоятельно вырабатывают ритмически повторяющиеся импульсы для тех или иных действий волокон.
Биохимические механизмы мышечной работы
В работе мышц участвует целая группа химических элементов, среди которых кальций и сократительные белки наподобие тропонина и тропомиозина. На базе этого энергетического обеспечения и выполняются рассмотренные выше физиологические процессы. Источником же этих элементов выступает аденозинтрифосфорная кислота (АТФ), а также ее гидролиз. При этом запас АТФ в мышце способен обеспечивать сокращение мышцы лишь в течение доли секунды. Несмотря на это, волокна могут отвечать на нервные импульсы в постоянном режиме.
Дело в том, что биохимические механизмы мышечного сокращения и расслабления с поддержкой АТФ связаны с процессом выработки резервного запаса макроэрга в виде креатинфосфата. Объем этого резерва в несколько раз превышает запас АТФ и в то же время способствует его генерации. Также помимо АТФ энергетическим источником для мышцы может выступать гликоген. К слову, на мышечные волокна приходится около 75% всего запаса данного вещества в организме.
Сопряжение возбудительных и сократительных процессов
В спокойном состоянии нити волокон не взаимодействуют друг с другом посредством скольжения, так как центры связок закрываются молекулами тропомиозина. Возбуждение может иметь место только после электромеханического сопряжения. Данный процесс также делится на несколько этапов:
- При активации нейромышечного синапса на мембране миофибриллы формируется так называемый постсинаптический потенциал, накапливающий энергию для действия.
- Возбуждающий импульс благодаря системе трубок расходится по мембране и активизирует ретикулум. Этот процесс в итоге способствует снятию барьеров с каналов мембраны, по которым выпускаются ионы, связывающиеся с тропонином.
- Белок тропонин, в свою очередь, открывает центры связок актина, после чего становится возможным механизм мышечных сокращений, но для его начала также потребуется соответствующий импульс.
- Использование открывшихся центров начнется в момент, когда к ним присоединятся головки миозина по описанной выше модели.
Полный цикл этих операций происходит в среднем за 15 мс. Период от начальной точки возбуждения волокон до полного сокращения называется латентным.
Процесс расслабления скелетной мышцы
При расслаблении мышц происходит обратный перенос ионов Са++ с подключением ретикулума и кальциевых каналов. В процессе выхода ионов из цитоплазмы количество центров связки сокращается, в результате чего происходит разъединение актиновых и миозиновых филаментов. Иными словами, механизмы мышечного сокращения и расслабления подключают те же функциональные элементы, но оперируют ими разными способами. После расслабления может наступать процесс контрактуры, при котором отмечается устойчивое сокращение мышечных волокон. Это состояние может сохраняться до момента, пока не наступит очередное действие раздражающего импульса. Бывает и контрактура краткого действия, предпосылками для которой становится тетаническое сокращение в условиях скопления ионов с большими объемами.

Фазы сокращения
Когда мускулатура приводится в действие раздражающим импульсом сверхпороговой силы, происходит одиночное сокращение, в котором можно выделить 3 фазы:
- Уже упомянутый выше период сокращения латентного типа, в процессе которого волокна накапливают энергию для совершения последующих действий. В это время проходят процессы электромеханического сопряжения и открываются центры связок. На данной стадии подготавливается механизм сокращения мышечного волокна, который активизируется после распространения соответствующего импульса.
- Фаза укорочения – длится 50 мс в среднем.
- Фаза расслабления – также длится примерно 50 мс.
Режимы мышечного сокращения
Работа при одиночном сокращении была рассмотрена как пример «чистой» механики мышечных волокон. Однако в естественных условиях такая работа не совершается, поскольку волокна находятся в постоянном отклике на сигналы двигательных нервов. Другое дело, что в зависимости от характера этого отклика может происходить работа в следующих режимах:
- Сокращения возникают при пониженной частоте импульсов. Если электрический импульс распространяется после завершения расслабления, то следует серия одиночных актов сокращения.
- Высокая частота импульсных сигналов может совпадать с расслабляющей фазой предшествующего цикла. В этом случае амплитуда, в которой работал механизм сокращения мышечной ткани, будет суммироваться, что обеспечит длительное сокращение с неполными актами расслабления.
- В условиях повышения частоты импульсов новые сигналы будут действовать в периоды укорочения, что спровоцирует длительное сокращение, которое не будет прерываться расслаблениями.
Оптимум и пессимум частоты
Амплитуды сокращений определяются частотой импульсов, которые раздражают мышечные волокна. В этой системе взаимодействия сигналов и откликов можно выделить оптимум и пессимум частоты. Первым обозначается частота, которая в момент действия будет накладываться на фазу повышенной возбудимости. В таком режиме может активизироваться механизм сокращения мышечного волокна с большой амплитудой. В свою очередь, пессимум определяет более высокую частоту, импульс которой приходится на фазу рефрактерности. Соответственно, в этом случае амплитуда уменьшается.
Виды работы скелетной мышцы
Мышечные волокна могут осуществлять работу динамически, статически и динамически-уступающе. Стандартная динамическая работа является преодолевающей – то есть мышца в момент сокращения перемещает объекты или его составные части в пространстве. Статическое действие мышцы в некотором роде избавлено от нагрузок, поскольку в этом случае не предусматривается изменение его состояния. Динамически-уступающий механизм мышечного сокращения скелетной мышцы срабатывает, когда волокна функционируют в условиях растяжения. Потребность в параллельном растяжении также может быть обусловлена тем, что работа волокон предполагает выполнение операций со сторонними телами.
В заключение
Процессы организации мышечного действия подключают самые разные функциональные элементы и системы. В работе задействуется сложный комплекс участников, каждый из которых выполняет свою задачу. Можно видеть, как в процессе активации механизма мышечных сокращений срабатывают и косвенные функциональные блоки. Например, это касается процессов генерации энергетического потенциала для совершения работы или системы блокировки центров связок, через которые происходит соединение миозинов и актинов.
Основная же нагрузка приходится непосредственно на волокна, которые выполняют те или иные действия по командам двигательных единиц. Причем характер выполнения определенной работы может быть разным. На него будут влиять параметры направляемого импульса, а также текущее состояние мышцы.
Энергия в мышцах освобождается при. Механизм мышечных сокращений. Функции и свойства скелетных мышц. Одиночное и тетанические сокращения
 в
в
Рисунок. Механизм сокращения: а – состояние покоя; б – умеренное сокращение; в – максимальное сокращение
Энергию для этого процесса поставляет гидролиз АТФ. Когда АТФ присоединяется к головке молекулы миозина, где локализован активный центр миозиновой АТФазы, связи между тонкой и толстой нитями не образуется. Появившийся катион кальция нейтрализует отрицательный заряд АТФ, способствуя сближению с активным центром миозиновой АТФазы. В результате происходит фосфорилирование миозина, т. е. миозин заряжается энергией, которая используется для образования спайки с актином и для продвижения тонкой нити. После того как тонкая нить продвинется на один «шаг», АДФ и фосфорная кислота отщепляются от актомиозинового комплекса. Затем к миозиновой головке присоединяется новая молекула АТФ, и весь процесс повторяется со следующей головкой молекулы миозина.
Затрата АТФ необходима и для расслабления мышц. После прекращения действия двигательного импульса Са 2+ переходит в цистерны саркоплазматического ретикулума. Тн-С теряет связанный с ним кальций, следствием этого являются конформаци-онные сдвиги в комплексе тропонин-тропомиозин, и Тн-I снова закрывает активные центры актина, делая их неспособными взаимодействовать с миозином. Концентрация Са 2+ в области сократительных белков становится ниже пороговой, и мышечные волокна теряют способность образовывать актомиозин.
В этих условиях эластические силы стромы, деформированной в момент сокращения, берут верх, и мышца расслабляется. При этом тонкие нити извлекаются из пространства между толстыми нитями диска А, зона Н и диск I приобретают первоначальную длину, линии Z отдаляются друг от друга на прежнее расстояние. Мышца становится тоньше и длиннее.
Скорость гидролиза АТФ при мышечной работе огромна: до 10 мк моль на 1 г мышцы за 1 мин. Общие запасы АТФ невелики, поэтому для обеспечения нормальной работы мышц АТФ должна восстанавливаться с той же скоростью, с какой она расходуется.
Расслабление мышцы происходит после прекращения поступления длительного нервного импульса. При этом проницаемость стенки цистерн саркоплазматической сети уменьшается, и ионы кальция под действием кальциевого насоса, используя энергию АТФ, уходят в цистерны. Удаление ионов кальция в цистерны ретикулума после прекращения двигательного импульса требует значительных энерготрат. Так как удаление ионов кальция происходит в сторону более высокой концетрации, т.е. против осмотического градиента, то на удаление каждого иона кальция затрачивается две молекулы АТФ. Концентрация ионов кальция в саркоплазме быстро снижается до исходного уровня. Белки вновь приобретают конформацию характерную для состояния покоя.
Таким образом, и процесс мышечного сокращения и процесс мышечного расслабления – это активные процессы, идущие с затратами энергии в виде молекул АТФ,
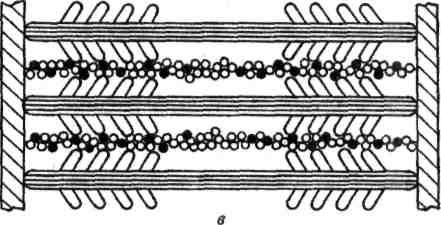
В гладких мышцах нет миофибрилл, которые состоят из нескольких сотен саркомеров. Тонкие нити присоединяются к сарколемме, толстые находятся внутри волокон. Ионы кальция также играют роль в сокращении, но поступают в мышцу не из цистерн, а из внеклеточного вещества, поскольку в гладких мышцах отсутствуют цистерны с ионами калькия. Этот процесс медленный и поэтому медленно работают гладкие мышцы.
—

—

—

—

—



Рисунок. Схема расположения толстых и тонких нией в гладких мышечных волокнах.
Источники:
http://www.bio.bsu.by/phha/06/06_text.html
http://www.syl.ru/article/340143/mehanizm-myishechnyih-sokrascheniy-kratko
http://studfile.net/preview/5920458/page:3/

